
Revised descriptions of New Zealand Cenozoic Mollusca from Beu and Maxwell (1990)

| | Revised descriptions of New Zealand Cenozoic Mollusca from Beu and Maxwell (1990) |
|
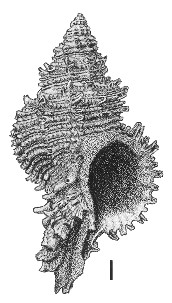  (Pl. 48l): GS4185, R22/f6502, Upper Castlecliff Shellbed, Castlecliff, Wanganui, Castlecliffian (GNS) |
Beu & Maxwell (1990): Chapter 16; p. 359; pl. 48 l.
Synonymy: Murex octogonus Quoy & Gaimard 1833, p. 531; Murex peruvianus Sowerby 1841 (in 1832-1841), pl. 195, fig. 103; Murexsul cuvierensis Finlay 1927a, p. 487; Muricopsis octogonus
Type species of Murexsul Iredale, 1915
Classification: Muricidae: Muricopsinae
Description: Moderately large for family (40-70 mm high), with moderately tall spire (about equal to aperture and canal), moderately long, open anterior canal, and large oval aperture, with lips smooth except for spinous digitations of the outer lip. Sculpture of many closely spaced, rounded, prominent, finely scaly spiral cords, each interspace filled by 1 narrow secondary cord; major cords raised into short to moderately long, backward- or upward-directed, semitubular spines where they cross thin, frilly varices; varices spaced regularly around whorls, 8-10 per whorl on large shells, producing roughly octagonal outline in plan view. Base of canal bears 2 or 3 rows of longer spines; fasciole very prominent, formed of a spiral row of many former anterior canals, producing a deep pseudumbilicus on large shells. Protoconch of 1.5 whorls, with flattened apex and margining keel, last half-whorl evenly convex.
Comparison: Marshall & Burch (2000) revised the Recent species and a few specimens of late Neogene fossil Murexsul, but most species in late Neogene rocks are still in need of revision. They established the new species M. profunda for specimens from 91-710 m off the Three Kings Islands, and M. (Rolandiella) scotti n. sp. for specimens from 13-168 m off eastern Northland, from the Three Kings Islands to Coromandel Peninsula. The subgenus M. (Rolandiella) is characterised by the shoulder spiral cord on early teleoconch whorls being medial on the whorl, and then ascending, and the intermediate spiral cords developes later than the others. In contrast, all species of Murexsul (sensu stricto) have 3 equally prominent spiral cords commencing together on the first spire whorl. The possible application of this subgenus to the many fossil species from New Zealand has not been considered as yet, one of the many questions remaining with the taxonomy of the genus.
The most similar fossil species is Murexsul marwicki (Long Beach Shellbed, Clifden, Southland, late Altonian), which has the same pattern of spination as M. octogonus, with two coarsely spinose cords around the neck, but has weaker spiral cords and so shorter spines on the varices. Most other named fossils referred here may not be closely related. M. espinosus (Mangapanian-Nukumaruan; ? = M. dilucidus, Opoitian) differs from M. octogonus in its smaller size, its much weaker sculpture without obvious spines, and in having a smoothly rounded (not flattened and keeled) protoconch apex; it is moderately common in Nukumaruan siltstone in Hawke's Bay and Wairarapa. The living Northland intertidal species M. mariae is similar to M. espinosus, but has a smaller protoconch with a flattened apex as in M. octogonus (Marshall & Burch 2000, fig. 27, 28). M. mariae apparently occurs fossil with M. octogonus in the young (Haweran, oxygen isotope stage 5a) cover beds of Hauriri Terrace, at the mouth of Wairoa Stream, Waverley Beach, west of Wanganui, according to Fleming's (1953) identifications, but this needs checking. If correct, it would indicate that the temperature at Wanganui was significantly higher when the terrace cover was deposited than it is at present.
Ponder (1972b, p. 237) synonymised Murexsul Iredale, 1915 with Muricopsis Bucquoy, Dautzenberg & Dollfus, 1882, but most muricid taxonomists since have regarded these as distinct genera, and Radwin & D'Attilio (1976) placed them in distinct subfamilies (Murexsul in Muricinae, Muricopsis in Muricopsinae). Vokes (1971a) treated Murexsul and Muricopsis as distinct genera in her catalogue of the Muricidae, and has since (e.g., Vokes & Houart 1986, p. 86, 87; see particularly the statement by Vokes 1988, p. 36) consistently maintained them as distinct genera. We agree with this action. Merle & Hourt (2003) have since used the distinct genera Murexsul and Muricopsis as examples of the way in which analyses of the ontogeny of spiral cords in Muricidae can elicudate phylogeny; they included both genera in subfamily Muricopsinae.
Distribution: Nukumaruan?; Castlecliffian-Recent; Recent, New Zealand (types of M. octogonus, M. peruvianus, and M. cuvierensis). Uncommon at only a few Castlecliffian fossil localities (notably the basal shellbed member of Shakespeare Cliff Sand at Wanganui, but a few specimens occur in Pinnacle Sand, Upper Castlecliff Shellbed, and Tainui Shellbed) and its ancestry is obscure. M. octogonus is common subtidally at present (commonly dredged on the inner-mid shelf around the northeastern North Island and in Cook Strait) and occurs on intertidal rocks in a few localities in the north-eastern North Island, from Auckland to Doubtless Bay.
Cite this publication as: "A.G. Beu and J.I. Raine (2009). Revised
descriptions of New Zealand Cenozoic Mollusca from Beu and Maxwell (1990). GNS
Science miscellaneous series no. 27."
© GNS Science, 2009
ISBN
978-0-478-19705-1
ISSN 1177-2441
(Included with a PDF facsimile file
copy of New Zealand Geological Survey Paleontological Bulletin 58 in CD version
from: Publications Officer, GNS Science, P.O. Box 30368 Lower Hutt, New
Zealand)