
Revised descriptions of New Zealand Cenozoic Mollusca from Beu and Maxwell (1990)

| | Revised descriptions of New Zealand Cenozoic Mollusca from Beu and Maxwell (1990) |
|
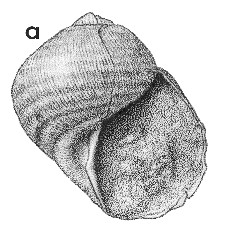  (Pl. 37a): Kaawa Creek, southwest Auckland, Opoitian (GNS, ex Fleming Collection) (apertural view of a slightly laterally compressed specimen). |
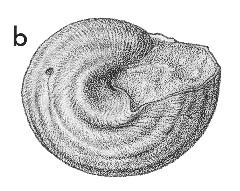 (Pl. 37b): Kaawa Creek, southwest Auckland, Opoitian (GNS, ex Fleming Collection) (basal view to show sinus). |
Beu & Maxwell (1990): Chapter 14; p. 292; pl. 37 a,b.
Synonymy: Hartungia typica Bronn 1861, p. 119; Janthina hartungi Mayer 1864, p. 242; Heligmope dennanti Tate 1893, p. 329; Turbo(?) postulatus Bartrum 1919, p. 100; Parajanthina japonica Tomida & Itoigawa 1982, p. 60; Bulbus (Heligmope) dennanti; Hartungia postulata
Type species of Hartungia Bronn, 1861 (= Heligmope Tate, 1893; = Parajanthina Tomida & Itoigawa, 1982; = Janthina Röding, 1798)
Classification: Janthinidae
Description: Large for family (17-40 mm high, 19-38 mm in diameter), shaped much as in the common "garden snail", Helix aspersa, with low, convex spire and large, enveloping, evenly inflated last whorl; imperforate; very thin and light, with a conspicuous pale brown outer calcific layer. Aperture very large, subcircular, with curved inner lip merging into strongly and regularly convex outer lip in apertural view; in basal view, a shallow sinus in base of outer lip slightly to right of junction with inner lip, producing a conspicuous costa parallel to inner lip on most specimens. Sculpture of low, indistinct, spiral folds in the shell wall, 9-11 on most specimens, almost as prominent on sutural ramp as lower down; whorl surface crossed by low, very thin, widely spaced, axial lamellae, about 1 mm apart on large specimens, Protoconch not seen, presumably pupiform and inclined to the teleoconch axis, as in Recent Janthina species.
Comparison: The succeeding species Janthina chavani (? = Hartungia chouberti Chavan 1951, p. 135) (Mangapanian-Nukumaruan in New Zealand) is very variable in spire height, large specimens having a much taller spire than adult J. typica, and its sutural ramp is less strongly convex than that of J. typica and lacks spiral folds. The presumed evolutionary change occurred abruptly in two widely separated New Zealand sequences (Te Ekaou Creek, Pohangina district, eastern Wanganui basin; Mohaka River, northern Hawke's Bay) at the Waipipian-Mangapanian boundary, suggesting this was an abrupt speciation event (punctated equilibrium) that produced a world-wide pelagic macrofossil datum-plane. Janthina chavani is common in the Roe Calcarenite, south-eastern Western Australia (Ludbrook 1978) and rare at a few localities in New Zealand (Mangapanian: Te Ekaou Creek, Pohangina; Mohaka River, shellbeds near base of sandstone zone upstream from rail viaduct; Nukumaruan: main highway near Utiku, Rangitikei Valley; Nukumaru Limestone, Nukumaru, Wanganui; Darkys Spur Formation, richly diverse fossiliferous sands halfway down Darkys Spur road, west of Devil's Elbow, central Hawke's Bay, mid-Nukumaruan). J. chavani seems little different from Hartungia chouberti, from Morocco, but there appear to be no other Northern Hemisphere records of this latest Pliocene form.
This species is a member of the Janthinidae, Recent species of which are cosmopolitan. Recently recognised evidence of a species intermediate between Hartungia and Janthina, without the spiral folds of J. typica and J. chavani, in late Pliocene rocks of Santa Maria Island in the Azores, suggests that this is all one clade. Hartungia seems to have been based on early species of Janthina. These shells float at the surface by means of a bubble raft, feeding on siphonophores, and casting ashore in persistent on-shore winds. The recorded localities for fossils appear to include most of the known Pliocene localities of sufficiently oceanic, wind-swept situation to have had janthinids blown ashore. The simplest scenario for the phylogeny of these taxa suggests that Hartungia is congeneric with Janthina, the strongly distinct, taller, unsculptured, brown janthinid Recluzia represents a clade distinct from Janthina, and Janthina and Recluzia developed the floating habit independently from distinct genera of Epitoniidae. If correct, this indicates that Janthinidae is part of Epitoniidae.
Distribution: Kapitean-Waipipian; Santa Maria Island, Azores Islands, eastern Atlantic, early-mid Pliocene (types of Hartungia typica and of Janthina hartungi); MacDonald's Bank, Muddy Greek, western Victoria, Australia, Kalimnan (type of Heligmope dennanti); Kaawa Creek, southwest Auckland, Opoitian (type of Turbo(?) postulatus); Higashigumi, near Mori-machi, Shizuoka Prefecture, Japan, Pliocene (type of Parajanthina japonica); common at a few localities in south-eastern Australia, near Perth in western Australia, at a few localities in north-eastern New Zealand, and rare at Santa Maria Island in the Azores and in the Canary Islands, eastern Atlantic; rare in Japan; not reported from elsewhere but presumably formerly cosmopolitan in tropical and temperate seas. In New Zealand, Janthina typica is rare in a few Kapitean localities near East Cape, relatively common at Kaawa Creek, southwest Auckland (Opoitian) and rare in other Opoitian localities in northern Hawke's Bay; and uncommon but widespread in Waipipian localities along the Wanganui-South Taranaki coast, in northern Hawke's Bay, and in Westland; presumably most are near-shore localities where on-shore winds have cast the pelagic snails onto beaches, as happens now around New Zealand with Janthina.
Cite this publication as: "A.G. Beu and J.I. Raine (2009). Revised
descriptions of New Zealand Cenozoic Mollusca from Beu and Maxwell (1990). GNS
Science miscellaneous series no. 27."
© GNS Science, 2009
ISBN
978-0-478-19705-1
ISSN 1177-2441
(Included with a PDF facsimile file
copy of New Zealand Geological Survey Paleontological Bulletin 58 in CD version
from: Publications Officer, GNS Science, P.O. Box 30368 Lower Hutt, New
Zealand)