
Revised descriptions of New Zealand Cenozoic Mollusca from Beu and Maxwell (1990)

| | Revised descriptions of New Zealand Cenozoic Mollusca from Beu and Maxwell (1990) |
|
  (Pl. 35d): holotype of Z. flemingi, GS3528, R11/f7014, Otahuhu Brewery well, Auckland, Waipipian (TM1369, GNS) |
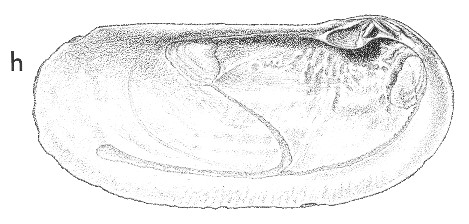 (Pl. 35h): holotype of Z. flemingi, GS3528, R11/f7014, Otahuhu Brewery well, Auckland, Waipipian (TM1369, GNS) |
Beu & Maxwell (1990): Chapter 14; p. 286; pl. 35 d,h.
Synonymy: Zenatia flemingi Marwick 1948, p. 23
Classification: Mactridae: Lutrariinae
Description: Large for genus (105 mm long), low and elongate, with very short anterior end and very long posterior end (i.e., umbo at anterior fifth), weakly inflated, with parallel, weakly down-curved, dorsal and ventral margins, regularly rounded anterior end, and rounded but slightly backward-sloping posterior end. Lunule a small, steeply sloping, striated area inside valve margin, immediately in front of hinge. Exterior sculptured only with weak growth ridges and weak periostracal wrinkles. Hinge with large bilobed cardinal tooth in left valve and pair of inverted-v- shaped cardinal teeth in right valve, margined by low anterior lateral tooth alongside lunule and very thin cardinal lamella alongside resilifer in left valve; a large, triangular resilifer below hinge plate behind teeth in both valves. Adductor scars large, subequal; pedal retractor scars small, separate, above anterior adductor; pallial sinus very deep, extending more than half length of shell.
Comparison: In living Zenatia acinaces, both anterior and posterior ends gape slightly, and dorsal and ventral margins are not parallel, allowing the animal to "rock" the valves open and closed during burrowing. Large specimens live in less than about 10 m of water, and smaller shells (to about 60 mm long) live in about 10 to 40 m. Zenatia is an endemic New Zealand genus of deep-burrowing mactrids of unusual long, narrow shape, still more extremely elongated than Lutraria (Pl. 39j, k). The Australian genus Zenatiopsis is still longer and narrower, with differently arranged pedal retractor muscle scars and a large internal strengthening ridge beneath the hinge; Zenatina is another Australian genus with a much more nearly central umbo, situated at the anterior third of the length.
A brief review of Zenatia species is in order, as a change is needed to the taxonomy of Beu & Maxwell (1990, p. 288), and a return to the conclusions by Beu (1966).
(1) The earliest species is Zenatia cretacea, based on a single small (length 44 mm) composite mould (internal mould of the hinge area and anterior end, external mould of the posterior end). Beu (1966) concluded that this specimen was from a Tertiary (Early-Middle Miocene?) concretion at Haumuri Bluff, but Warren & Speden (1977, p. 45, table 2, fig. 27) regarded the holotype of Z. cretacea as a Piripauan (Late Cretaceous) specimen from the base of the Okarahia Sandstone at Haumuri Bluff, Marlborough. However, no further Cretaceous specimens have been collected, and a recent dinoflagellate and pollen sample from the holotype of another supposedly Cretaceous species form Haumuri Bluff, Maoricrypta hochstetteriana (Wilckens 1922, p. 5, fig. 9a, b), contained Acacia and is at most Middle Miocene in age (G. Wilson, formerly of GNS, pers. comm.), demonstrating that A. McKay collected a significant number of Miocene fossils from concretions at Haumuri Bluff, and any suspect taxa that have not been re-collected from Cretaceous rocks are likely to be Cenozoic. We therefore return to the earlier concept that the holotype of Z. cretacea is a Miocene specimen, and this name applies to the earliest Cenozoic species of Zenatia. This species (Whaingaroan-Altonian; possibly also Altonian-Waiauan) is small (rarely more than 60 mm long), with the lunule sloping outward steeply; the cardinal angle (angle between the cardinal teeth of the right valve, or between the arms of the cardinal tooth of the left valve) averages 73°. Specimens of Altonian-Waiauan age are poor, but appear to be intermediate between the earlier Zenatia cretacea and the later Z. acinaces, i.e., this change appears to have been anagenetic.
(2) Z. acinaces (Tongaporutuan-Recent): large (shallow-water specimens to 100 mm long), cardinal angle averages 71°, lunule the shortest and widest of the genus, flat on the hinge plate or sloping slightly inwards.
(3) Z. flemingi (Waipipian, Otahuhu well only): large, cardinal angle averages 81°, lunule slightly longer, narrower, and more outward-sloping than in Z. acinaces.
(4) Z. tepikiensis (Haweran, Te Piki member, near East Cape, only): status uncertain as known by immature specimens only (to 53 mm long); cardinal angle averages 88°, i.e., this form appears to belong in a "northern lineage" and to be descended from Z. flemingi rather than to be related to Z. acinaces; but a close relationship to Z. acinaces is possible.
(5) Z. waipipiana (Waipipian, Wanganui-South Taranaki coast only): the subgenus Z. (Zenatraria) was proposed by Beu (1966) for species with a rounded (rather than obliquely angled) posterior outline, a narrow cardinal angle (averaging 55° in both species formerly assigned here), a long, rather narrow, bevelling lunule sloping outward towards the dorsum, and differently arranged pedal retractor muscle scars from those of Zenatia (Zenatia). However, "Z". vellai has proven to belong in Lutraria, and the subgenus is not recognised. Z. waipipiana has a very short anterior end, with a steeply angled lunule, and the lower pedal retractor muscle scar is elongate, subrectangular, and just above the top of the anterior adductor muscle scar, closer to it than in Z. acinaces.
(6) "Zenatia (Zenatraria)" vellai (Nukumaruan, Wanganui basin and southern Wairarapa; uncommon in Nukumaru Brown Sand, but rare elsewhere) proved to belong in Lutraria (Psammophila) (Beu 2006, p. 254), demonstrating that Zenatia is little different from and presumably closely related to Lutraria, and that a subfamily Zenatiinae is superfluous. L. vellai is more inflated than Zenatia species, with a relatively long anterior end whose dorsal outline is a little straightened by the large, bevelling lunule; the lower pedal retractor muscle scar is an oval facet on the posterior edge of the anterior adductor muscle scar, very similar to that in the Recent Japanese species L. (Psammophila) sieboldtii Deshayes in Reeve, 1854 (Beu 2006). However, it is also much smaller and more weakly inflated than the large, robust species it occurs alongside, Lutraraia (Lutraria) grandis (Hutton, 1873).
Distribution: Waipipian, Otahuhu Brewery well, Auckland, Waipipian (type); poor moulds from a few nearby surface exposures, otherwise not known elsewhere; but very similar to the Tongaporutuan-Recent Z. acinaces.
Cite this publication as: "A.G. Beu and J.I. Raine (2009). Revised
descriptions of New Zealand Cenozoic Mollusca from Beu and Maxwell (1990). GNS
Science miscellaneous series no. 27."
© GNS Science, 2009
ISBN
978-0-478-19705-1
ISSN 1177-2441
(Included with a PDF facsimile file
copy of New Zealand Geological Survey Paleontological Bulletin 58 in CD version
from: Publications Officer, GNS Science, P.O. Box 30368 Lower Hutt, New
Zealand)